Tato kapitola obsahuje popis životního cyklu kapradin a jejich rozmanitých odchylek.
Seznam kapitol:
1. Přehled životního cyklu:1. Základní přehled
2. Spory:
2.0 Úvod | 2.1 Dělení spor dle životaschopnosti - abortace | 2.2 Dělení spor dle barvy - přítomnosti chlorofylu
1. Přehled životního cyklu
1. Základní přehled
Tato kapitola slouží pro začátečníky, ukazuje základní prvky životního cyklu vyšších rostlin, přesněji kapradin.
Kapradiny patří mezi vyšší rostliny, tedy mechorosty, kapraďorosty a semenné rostliny. I když všechny tyto skupiny mají své odlišnosti v rozmnožování, mají v základu stejný životní cykus. Střídají se při něm dvě fáze lišící se ploidní úrovní, zjednodušeně počtem chromozomů (nositelů genetické informace) v jádře buněk. Haploidní generace je taková, která má jednu sadu chromozomů, označuje se "n". Diploidní fáze má sady dvě, označuje se "2n".
Toto rozčlenění generací je důležité pro všechny sexuální organismy, spojením dvou jednotlivých (haploidních) sádek totiž vzniká jedinec s vlastnostmi získanými od obou rodičů, to posiluje rozmanitost potomků. Jaká je tedy situace u rostlin?
Haploidní ("n") fáze u rostlin se nazívá "gametofyt", tvoří totiž gamety, pohlavní buňky. Diploidní fáze ("2n") se nazývá sporofyt, tvoří totiž spory, jeho první buňka (vzniklá splynutím dvou gamet) se nazývá zygota. Zjednodušeně se cyklus dá vyjádřit takto:
Gametofyty jsou u kapradin drobné přehližené rostlinky o velikosti v milimetrech až centimetrech. Tvoří dva typy rozmnožovacích útvarů - gametangií - tvořící pohlavní buňky - gamety: a) Archegonia tvoří vajíčka. b) Antheridia tvoří spermatozoidy. Mnohobičíkaté spermatozoidy musí doplavat k archegoniu a prolézt dovnitř, následně dochází k oplození a vzniku zygoty, ve které se obě gamety (n) spojily dohromady, zygota je tedy diploidní (2n) a začíná fáze sporofytická.
Sporofyty u kapradin jsou často tvořeny velikými listy rostoucími z podzemního stonku, oddenku. Sporofyty jsou to, co si většina z nás představí pod pojmem kapradina. Rozmnožovací útvary sporofytu se nachází nejčastěji na spodní straně listů a nazývají se sporangia (výtrusnice). Často se shlukují do kupek, které mohou být chráněné ochrannou blankou, ostěrou. Ve sporangiích je skupinka buněk, u kterých dochází k redukčnímu dělení, meióze. Výsledné dceřinné buňky tohoto dělení mají jen polovinu chromozomů, proto redukční dělení (2n -> n). Tyto dceřinné buňky se obalí ochrannými obaly a vylétávají ze sporangií, nazýváme je spory. Klíčí do gametofytů, tím se cyklus uzavírá.
Tradičně zmiňovanou výjimku tvoří přesličky. I přes úplně jiný tvar sporofytu respektují některé přesličky výše zmíněný systém životního cyklu. Ale například u běžné přesličky rolní (Equisetum arvense) má sporofyt navíc systém dvou typů lodyhy. Na jaře vyrůstá z podzemního oddenku béžová jarní lodyha. Nemá fotosyntetická barviva a žije jen ze zásob z oddenku. Po vytvoření spor jarní lodyha umírá a ze stejného oddenku roste zelená letní lodyha. Tato lodyha normálně fotosyntetizuje a tvoří zásoby na další jaro. Gametofytická fáze je totožná, jako u ostatních kapradin. Ze zygoty vyrůstá zelená lodyha.
Tento systém jarní "předčasné" tvorby spor umožňuje přesličce využít časové okno na začátku roku, kdy odhalenou půdu ještě nestihli zarůst silnější konkurenti.
{kind=link}
{kind=link}
2. Spory
2.0 Úvod
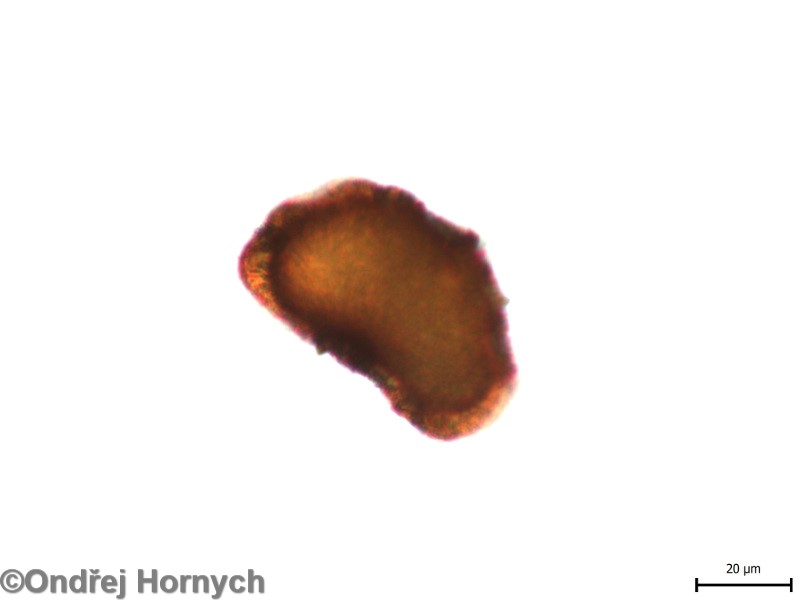
Obr. 1: Kapraď Borrerova - Dryopteris borreri - monoletní spora s tmavším ledvinitým exosporem a světlejším okolním perisporem.
Spory jsou jednobuněčné, drobné, většinou haploidní rozmnožovací útvary rostlin. Vznikají meioticky, redukčním dělením, ve sporangiích. Jedná se o první buňku gametofytní generace. U apogamních druhů mají spory stejnou ploidní úroveň, jako sporofyty, přestože většinou vznikají meiózou. Více viz apogamie. Těmto sporám se říká diplospory.
Spora je buňka chráněná vnějším vícevrstevným obalem - sporodermem. Od středu první vrstva, endospor je převážně tvořená celulózou a odděluje ostatní vrstvy od plasmatické membrány a tím obsahu buňky. Tato vrstva není pozorovatelná v mikroskopu. Prostřední vrstva, exospor, je hladká a udává tvar spory. Perispor je vnější vrstva sporodermu, často bývá ornamentovaná, například zvlněná, nebo ostnitá. Exospor a perispor jsou tvořeny z velmi odolného materiálu sporopoleninu a jejich tvorba je zajišťovaná sporangiem, ne sporou samotnou. Jsou dobře pozorovatelné pod mikroskopem. Detailní popisy spor různých rodů kapradin jsou k nalezení v Tryon et Lugardon 1990.
Spory kapradin lze dle tvaru a typu apertury (místo, kde je obal spory tenčí, umožňující klíčení) rozdělit do dvou kategorií – triletní a monoletní. Triletní spory jsou +- radiálně symetrické, mají tvar čtyřstěnu. Apertura je trojitá, tři stejná ramena apertury vybíhají ze společného středu. Monoletní spory jsou častější, než triletní. Jsou bilaterálně symetrické a mají často ledvinitý tvar (Obr. 1). Apertura je tvořená jednou čarou.
Některé kapradiny jsou známé arsenové bioakumulátory, vytahují z půdy jedovatý arsen a ukládají ho do svých tkání. Mezi typické příklady patří Pteris vittata, která snese 200x vyšší množství arsenu, než například jiná kapradina, Ceratopteris richardii (Gumaelius et al. 2004). P. vittata navíc dokáže vytáhnout velké množství arsenu ze zamořených i nezamořených půd a ukládat ho do spor. Jedna rostlina dokáže tímto způsobem ročně mobilizovat až 7,8 mg arsenu do spor a tím do ovzduší i z arsenem nezamořených půd (Lessl et al. 2015).
Potenciálně karcinogenní účinky mohou mít spory různých kapradin, například hasivky (Pteridium aquilinum). Extrakt spor způsobuje rozpad DNA, čímž může podporovat vznik rakoviny, lidská játra jsou schopná tuto rakovinotvornost téměř úplně odstranit (Simán et al. 2000).
Shrnutí klíčových informací:
- Spory jsou jednobuněčné rozmnožovací útvary rostlin. Vznikají ve sporangiích a klíčí v gametofyt.
- Spory vznikají meiozou, s výjimkou apomiktických taxonů jsou tedy haploidní.
- Spory jsou obaleny několikavrstevným ochranným obalem - sporodermem, složeným převážně ze sporopoleninu.
- Spory lze dělit dle tvaru na triletní a monoletní.
2.1 Dělení spor dle životaschopnosti – abortace

Obr. 2: Kapraď - Dryopteris ×deweveri - tmavá abortovaná spora s narušenou ornamentací.
Pokud proběhne proces sporogeneze, tvorby spor, v pořádku, ve sporangiu vznikají životaschopné spory. Tvar, velikost a barva výsledné spory odpovídá danému druhu a za správných podmínek je spora schopná klíčit v gametofyt. Kromě životaschopných spor ale každá rostlina produkuje některé spory vyvinutých nesprávně, abortované. Tyto abortované spory jsou neschopné klíčení a pro rostlinu nemají žádný další význam.
Abortované spory se ne vždy snadno poznají, ale většinou bývají výrazně tmavší, než spory vyvinuté (Obr. 2). Jejich exospora je zkolabovaná a nemá správný tvar (například ledvinitý u monoletních druhů). Abortované spory mají vysokou variabilitu ve velikosti, od drobného prachu po několikanásobně velké koule. Pro určování abortace je k nalezení je více informací viz Wagner et Chen 1965, nebo Hornych et Ekrt 2017.
Existuje několik důvodů, kapradiny tvoří abortované spory. Předpokladem pro řádnou tvorbu spor je správné spárování chromosomů během meiózy. Pokud při tomto procesu nastane chyba, nebo není možné chromosomy spárovat, výsledná spora je vždy abortovaná. Tento proces je dobře pozorovatelný u hybridů, kteří mají dvě odlišné sady chromosomů od dvou různých druhů. Hybridi proto tvoří téměř všechny spory abortované. Tento jev je u kapradin téměř univerzální a kontrola abortace spor je hlavní určovací znak odlišující hybridy od jejich rodičovských druhů (Wagner et Chen 1965). Jedinou výjimku z tohoto pravidla tvoří apomiktiční hybridi, viz níže.
Mimo genetické bariéry pro tvorbu životaschopných spor může mít na správnou sporogenezi vliv i okolní prostředí, či stav rostliny. Sporogeneze je energeticky velmi náročný proces. I když neexistují přímé doklady, nevhodné životní prostředí a okolní vlivy zcela jistě snižují schopnost tvorby životaschopných spor. I životní historie jedince může sporogenezi ovlivnit. Například je znám případ, kdy sleziník Asplenium aethiopicum vyprodukoval nadstandardně velké množství spor jeden rok a druhý rok tvořil, zřejmě z vyčerpání, většinu spor abortovaných (Braithwaite 1964).
Ať už je důvod jakýkoli, každá rostlina tvoří určité procento abortovaných spor. Jak již bylo zmíněno, hybridi tvoří vyvinuté spory výjimečně a procento abortace je málokdy nižší, než 99%. Naopak normální sexuálně se rozmnožující druhy tvoří průměrně 3-5% abortovaných spor (Gomes et al. 2006; Quintanilla et Escudero 2006; Arosa et al. 2009; Hornych et Ekrt 2017). Jsou ale známy i případy kdy jedinci tvořili téměř pětinu spor abortovaných (Quintanilla et Escudero 2006; Hornych et Ekrt 2017).
Apomixie, nepohlavní tvorba sporofytu bez spojení gamet, může výrazně ovlivnit schopnost druhu tvořit správně vyvinuté spory. U rodu Dryopteris a Phegopteris tvoří apomikti průměrně asi 18% abortovaných spor (Hornych et Ekrt 2017). Zvýšené procento abortace bylo pozorováno i u Cornopteris christenseniana (Park et Kato 2003), nebo rodu Pteris (Walker 1962). Někteří autoři ale přichází s odlišnými výsledky naznačujícími procento abortace bližší sexuálním druhům, například pro rod Cyrtogonellum (Guo et Liu 2013), Pteris vittata (Khare et Kaur 1983), nebo Dryopteris affinis (Quintanilla et Escudero 2006).
Na závěr stojí za zmínku speciální případ, který si nedávno získal pozornost vědců, apomiktičtí hybridi. Většina hybridů kapradin má oba rodiče sexuálně se rozmnožující, ale apomiktičtí hybridi mají vždy otcovský druh rozmnožující se apomikticky. Takové hybridní sporofyty jsou potom schopni tvořit 5-20% vyvinutých spor (Eschelmüller 1998; Fraser-Jenkins 2007; Ekrt et Koutecký 2016; Hornych et Ekrt 2017). Tento fakt je výrazně odlišuje od běžných hybridů a umožňuje jim zpětné křížení a tvorbu dalšího hybridního potomstva (Ekrt et Koutecký 2016).
Shrnutí klíčových informací:
- Některé spory kapradin neprojdou správným vývojem a jsou nefunkční – abortované.
- Abortace je ovlivněna vnitřními i vnějšími podmínkami.
- Většina hybridů kapradin se dá dobře poznat díky téměř 100% abortaci spor.
- Sexuálně se rozmnožující druhy mají většinou míru abortace velmi nízkou.
- Apomiktické druhy tvoří průměrně téměř pětinu spor abortovaných, ale tento trend neplatí pro všechny případy.
2.2 Dělení spor dle barvy - přítomnosti chlorofylu
Spory různých druhů kapradin mají rozličné barvy, od téměř bezbarvých spor Phegopteris connectilis, přes sytě žluté spory Athyrium filix-femina, po tmavě hnědé spory Polystichum aculeatum. Úplně abortované spory mohou mít barvu až téměř úplně černou.
Tradičně se ale druhy dělí, dle barvy spor, na druhy se sporami zelenými a nezelenými.
Taxonů se zelenými sporami je menšina. V České republice tvoří zelené spory zhruba pětina druhů. Patří mezi ně přesličky (Equisetum spp.), podezřeň královská (Osmunda regalis), pérovník pštrosí (Matteuccia struthiopteris) a vláskatec tajemný (Hymenophyllum speciosum), který u nás ale spory netvoří. Po světě existují další druhy, například celé čeledi Hymenophyllaceae, Onocleaceae a Osmundaceae, nebo někteří zástupci čeledi Polypodiaceae (např. podčeleď Grammitidae, nebo Marginariopsis wiesbaurii). Jednotlivé druhy se vyskytují i v čeledích Blechnaceae, Lomariopsidaceae a Pteridaceae (Sundue et al. 2011). Tento seznam nejspíše není kompletní a časem se najdou další druhy.
Zelenou barvu spor způsobuje přítomnost chloroplastů plných chlorofylu. Brzy se zjistilo, že tyto spory mají kromě odlišné barvy i jiné výrazné rozdíly ve srovnání se sporami nezelenými shrnuté v Tab. 1.
Tab. 1: Srovnání druhů se zelenými a nezelenými sporami, tučně jsou zvýrazněny výhodnější hodnoty pro danou vlastnost. Data upravena z Lloyd et Klekowski (1970).
| Vlastnost | Druhy se zelenými sporami | Druhy s nezelenými sporami |
| Přítomnost chlorofylu ve spoře | ano | ne |
| Počty druhů | stovky | tisíce až desetitisíce |
| Rychlost růstu gametofytů | velmi rychlá, nebo pomalá | rychlá, nebo pomalá |
| Průměrná doba do vyklíčení | 1,46 dní | 9,5 dní* |
| Průměrná životnost spor | 48 dní | 1045 dní (cca 2,8 let) |
Přítomnost chloroplastů a tím aktivního fotosyntetického aparátu může urychlit růst gametofytu, ale epifytické druhy se zelenými sporami často tvoří dlouhověké, pomalu rostoucí gametofyty (Lloyd et Klekowski 1970). Takové epifytické druhy (Polypodiaceae, podčeleď Grammitidae) mají z neznámého důvodu rychlejší evoluci chloroplastového genomu (Schneider et al. 2004).
Zelené spory klíčí rychleji, než spory nezelené, je u nich zrušena dormantní fáze. Zelené spory klíčí většinou do tří dnů (Lloyd et Klekowski 1970), na rozdíl od až 210 dní, které potřebuje nezelená spora druhu Plagiogyria glauca na vyklíčení (Stokey et Atkinson 1956). Rod Trichomanes to dotáhl tak daleko, že jeho zelené spory mohou klíčit již ve sporangiu a buňky se dělí již ve sporovém obalu (Stokey 1940). U homosporických kapradin se toto endosporické klíčení vyskytuje jen u druhů se zelenými sporami (Sundue et al. 2011).
Za tyto výhody platí zelené spory snížením doby životnosti - schopnosti klíčit – oproti metabolicky pasivním nezeleným sporám. Zatímco nezelené spory jsou schopny klíčit i po desítkách let spánku, zelené spory vydrží přežít pár dní, maximálně jeden rok (Lloyd et Klekowski 1970). Například u přesličky zimní (Equisetum hyemale) se ukázalo, že po dvou týdnech sucha nejsou spory schopny obnovit v dostatečné míře fotosyntézu, kvůli rozpadu fotosyntetického aparátu, a tím přicházejí o schopnost úspěšně vyklíčit. Pokud byly ale spory suché jen jeden den, po zvlhčení dosahují čistého fotosyntetického zisku již za šest minut (Lebkuecher 1997). Přítomnost perisporu (např. u Onocleaceae) může mít positivní vliv na dobu klíčivosti (Lloyd et Klekowski 1970)
Neschopnost zelených spor klíčit již po krátké době může být velký problém, pokud chceme spory uschovat na delší dobu, například do sporových bank, nebo na pozdější použití. Ballesteros et al. (2011) dokázali prodloužit dobu klíčivosti u Equisetum ramosissimum a Osmunda regalis teoreticky až na několik set let při uchování spor v tekutém dusíku. Tito autoři dále ukázali, že spory je třeba uložit velmi rychle po dozrání pro získání maximální doby klíčivosti.
Pomocí epiflorescenční mikroskopie se podařilo Sundue et al. (2011) detekovat chlorofyl i ve sporách tradičně udávaných, jako nezelené, mezi nimi byly například spory olivově zbarvené. Pro tento typ spor přišli s označením kryptochlorofylní, jelikož chlorofyl je přítomen, ale pod světelným mikroskopem není vidět, například skrze tmavý obal spory. Pátráním ve třech rodech kde se vyskytují zelené i nezelené spory se podařilo Sundue et al. (2011) najít 43 druhů s kryptochlorofylními sporami, například Elaphoglossum lehmannianum. Pod epiflorescenčním mikroskopem se navíc autorům podařilo detekovat zbytky chlorofylu i v zelených sporách starých 110 let.
Další vědecké práce o druzích s kryptochlorofylními sporami bohužel zatím nevznikly, ale lze předpokládat, že existují další takové druhy. Koncept kryptochlorofylních spor výrazně narušuje klasické pojetí spor dle barvy a možná by pomohl vysvětlit anomálie v chování některých nezelených spor, které se chovají „zeleně“.
Shrnutí klíčových informací:
- Spory lze dělit dle přítomnosti chlorofylu na zelené - chlorofylní a nezelené
- Zelené spory obsahují chlorofyl a mají aktivní chloroplasty, proto jsou schopné klíčit téměř okamžitě a rychle, jejich životnost je ale výrazně snížena.
- Nezelené spory chlorofyl neobsahují a jsou schopny klíčit až po několika týdnech, díky neaktivnímu metabolismu jsou schopné klíčít i po několika letech.
- Mezi významné české kapradiny se zelenými sporami patří například přesličky, nebo pérovník pštrosí. Zhruba pětina naší pteridoflóry tvoří zelené spory.
Zdroje
- Arosa M. L., Quintanilla L. G., Ramos J. A., Ceia R., Sampaio H. (2009): Spore maturation and release of two evergreen Macaronesian ferns, Culcita macrocarpa and Woodwardia radicans, along an altitudinal gradient. - Amer. Fern J. 99: 260–272. doi: 10.1640/0002-8444-99.4.260
- Ballesteros D., Estrelles E., Walters C. & Ibars A. M. (2011): Effect of storage temperature on green spore longevity for the ferns Equisetum ramosissimum and Osmunda regalis. – CryoLetters 32(2): 89-98.
- Braithwaite A. F. (1964): A New type of apogamy in ferns. - New Phytol. 63: 293–305.
- Ekrt L., Koutecký P. (2016): Between sexual and apomictic: unexpectedly variable sporogenesis and production of viable polyhaploids in the pentaploid fern of the Dryopteris affinis agg. (Dryopteridaceae). - Ann. Bot. 117: 97–106. doi: 10.1093/aob/mcv152
- Eschelmüller A. (1998): Keimversuche mit Sporen der triploiden Sippen von Dryopteris affinis und ihren Bastarden mit Dryopteris filix-mas. - Mitteilungen des Naturwissenschaftlichen Arbeitskreises Kempten 36: 47–78.
- Fraser-Jenkins C. R. (2007): The species and subspecies in the Dryopteris affinis group. - Fern Gaz. 18:1–26.
- Gomes S. G., Randi A. M., Puchalskil A., Santos D. D. S., dos Reis M. S. (2006): Variability in the germination of spores among and within natural populations of the endangered tree fern Dicksonia sellowiana Hook. (Xaxim). - Braz. Arch. Biol. Technol. 49: 1–10. doi: 10.1590/S1516-89132006000100001
- Gumaelius L., Lahner B., Salt D. & Banks J. A. (2004): Arsenic hyperaccumulation in gametophytes of P. vittata: a new model system for analysis of arsenic hyperaccumulation. – Plant Physiol. 136: 3198–3208.
- Guo Z. Y., Liu H. M. (2013): Gametophyte morphology and development of three species of Cyrtogonellum Ching (Dryopteridaceae). - Amer. Fern J. 103: 153–165. doi: 10.1640/0002-8444-103.3.153
- Khare P. B., Kaur S. (1983): Gametophyte differentiation of pentaploid Pteris vittata L. - Proc. Natn. Acad. Sci. India 49: 740–742.
- Lebkuecher J. G. (1997): Desiccation-time limits of photosynthetic recovery in Equisetum hyemale (Equisetaceae) spores. - Am. J Bot. 84(6): 792-797.
- Lessl J. T., Guan D. X., Sessa E., Rathinasabapathi Bala & Ma L. Q. (2015): Transfer of arsenic and phosphorus from soils to the fronds and spores of arsenic hyperaccumulator Pteris vittata and three non-hyperaccumulators. – Plant Soil 390: 49–60.
- Lloyd R. M. & Klekowski E. J. (1970): Spore Germination and Viability in Pteridophyta: Evolutionary Significance of Chlorophyllous Spores. – Biotropica 2(2): 129-137.
- Park C., Kato M. (2003): Apomixis in the interspecific triploid hybrid fern Cornopteris christenseniana (Woodsiaceae). - J. Pl. Res. 116: 93–103. doi:10.1007/s10265-003-0081-y
- Quintanilla L. G., Escudero A. (2006): Spore fitness components do not differ between diploid and allotetraploid species of Dryopteris (Dryopteridaceae). - Ann. Bot. 98: 609–618. doi: 10.1093/aob/mcl137
- Schneider H., Smith A. R., Cranfill R., Hildebrand T. J., Haufler C. H. & A. Ranker T. A. (2004): Unraveling the phylogeny of polygrammoid ferns (Polypodiaceae and Grammitidaceae): exploring aspects of the diversification of epiphytic plants. – Mol. Phylegenet. Evol. 31:1041-1063.
- Simán S. E., Povey A. C., Ward T. H., Margison G. P. & Sheffield E. (2000): Fern spore extracts can damage DNA. – Br. J Cancer 83(1): 69-73.
- Srivastava M., Ma L. Q., Rathinasabapathi B. & Srivastava P. (2009): Effects of selenium on arsenic uptake in arsenic hyperaccumulator P. vittata L. - Bioresour. Technol. 100: 1115–1121.
- Stokey A. G. (1940): Spore germination and vegetative stages of the gametophytes of Hymenophyllum and Trichomanes. - Bot. Gaz. 101: 759-790.
- Stokey A. G. & Atkinson L. R. (1956): The gametophytes of Plagiogyria glauca (Bl.) Mett. and P. semicordata (Pr.) Christ. - Phytomorphol. 6: 239-249.
- Sundue M., Vasco A. & Moran R. C. (2011): Cryptochlorophyllous Spores in Ferns: Nongreen Spores that Contain Chlorophyll – Int. J. Plant Sci. 172(9): 1110-1119.
- Tryon A. F. & Lugardon B. (1990): Spores of the Pteridophyta: surface, wall structure, and diversity based on electron microscope studies. - Springer, New York.
- Wagner W. H. Jr & Chen L. C. (1965): Abortion of spores and sporangia as a tool in the detection of Dryopteris hybrids. - Amer. Fern J. 55: 9–29.
- Walker T. G. (1962): Cytology and evolution in the fem genus Pteris L. - Evolution 16: 27–43.